ASSOCIATION (OU "CONCATENATION" OU "LINKAGE")
Jusqu'à présent nous avons traité le croisement mendélien dihybride (ou polyhybride) en supposant que les différentes paires alléliques se trouvent effectivement sur différentes paires de chromosomes homologues.Mais le nombre de paires chromosomiques, bien que différent d'une espèce à l'autre, varie dans des limites étroites (peu d'espèces atteindre une centaine de chromosomes), alors que le nombre de gènes se compte par dizaines de milliers.
Que les caractères choisis par Mendel pour ses expérimentations soient tous ségrégués indépendamment (sans donc brouiller les calculs dans la distribution des phénotypes dans le F2 du croisement polyhybride) était un heureux hasard.Si deux paires d'allèles étaient trouvées sur des loci adjacents, la loi qui s'ensuivrait s'appellerait la loi d'association.
Sachant que de très nombreux caractères ont leur locus sur un même chromosome et que ce sont les paires chromosomiques qui se séparent indépendamment dans la méiose, on comprend bien combien il arrive fréquemment que deux paires de caractères, si elles étaient associées sur un chromosome dans le parent l'organisme, restent associés également dans le gamète et donc dans l'organisme auquel il apportera son propre matériel génétique.
On voit ainsi que « l'association représente » une exception, loin d'être rare, à l'indépendance énoncée dans la troisième loi de Mendel.
ÉCHANGE OU « CROISEMENT » ET RECOMBINAISON
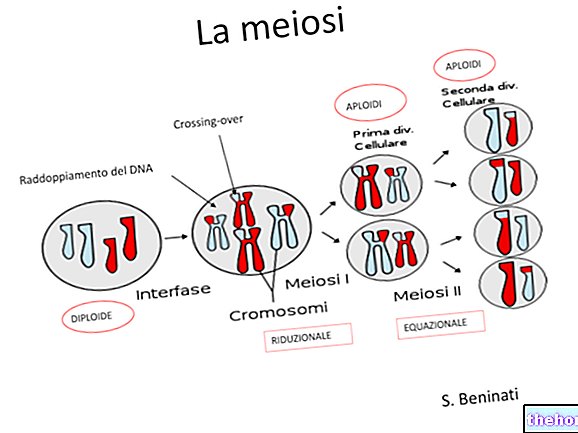
En parlant de méiose, nous avons indiqué qu'il existe deux moments différents de mélange du matériel génétique : l'un est celui de la ségrégation des chromosomes dans les gamètes, et c'est celui observé par Mendel.
L'autre moment, qui précède en fait, est celui où les quatre chromatides de chaque paire de chromosomes homologues s'échangent mutuellement des traits identiques, à la suite de cet échange, deux facteurs qui étaient associés sur un même chromosome seront au contraire indépendants dans les gamètes. qu'un échange ait lieu est proportionnel, en première approximation, à la longueur du chromosome, et dans les chromosomes plus longs il peut y avoir encore plus qu'un échange.
Le phénomène peut être détecté cytologiquement, en observant un nombre suffisant de méioses au microscope.
Le taux de re-match est le taux auquel deux caractères qui étaient associés dans la génération parentale se recombinent différemment dans F2.
Si les deux loci sont absolument contigus, la probabilité qu'un chiasma les sépare sera pratiquement nulle. Le taux de recombinaison sera de : n° recombinants. Si deux loci sont sur deux chromosomes différents, le taux de recombinaison sera de 0,5 (probabilité égale, pour deux caractères qui se sont joints à la génération P, de se retrouver aléatoirement ensemble en F2). Le taux de recombinaison peut donc varier entre 0,0 et 0,5. Pour les petites distances sur le chromosome, la distance et le taux de recombinaison sont directement proportionnels. Pour des distances plus longues, il est possible que deux échanges aient lieu entre deux loci. Il apparaît maintenant clair que deux facteurs séparés par deux échanges sont à nouveau associés. il est clair, à ce stade, que la proportionnalité entre la distance des loci et la probabilité de recombinaison est perdue.
Les loci trouvés associés sur le même chromosome constituent des «groupes d'association». Des loci très éloignés peuvent avoir une telle probabilité de séparation par échange qu'ils se comportent comme indépendants, mais chacun d'eux sera associé, avec un taux de recombinaison plus faible, aux loci intermédiaires.
Lorsque les taux de recombinaison entre de nombreuses paires de gènes au sein d'un groupe d'association sont connus, la construction de « cartes génétiques » peut commencer. En gardant à l'esprit que la distance entre deux gènes (a et b) s'exprime par le taux de recombinaison et que la distance de a à un troisième gène c peut être soit la somme soit la différence par rapport à sa distance à b, il est possible pour reconstruire une carte des distances réciproques, qui sera la carte génétique au sein de ce groupe d'association, c'est-à-dire de ce chromosome.
Nous devons maintenant considérer en général quelques concepts qui limitent la manifestation phénotypique des caractères génotypiques.
Nous parlerons tout d'abord des notions de pénétrance et d'expressivité, puis nous consacrerons une attention particulière aux phénomènes de régulation de l'action des gènes.
PÉNÉTRANCE
La pénétrance d'un gène représente sa capacité à se manifester dans le phénotype. La pénétrance est mesurée statistiquement en comptant la fréquence des phénotypes qui montrent ce caractère sur 100 génotypes qui le contiennent. Un trait avec une pénétrance de 0,7 est un trait qui se produit phénotypiquement dans 70 % de sa fréquence génotypique.
EXPRESSIVITÉ
L'expressivité est une évaluation quantitative du degré de manifestation phénotypique.
REGULATION DE L'ACTION DES GENE
Les cellules produisent toutes leurs enzymes et protéines à la même vitesse et en même temps. Les cellules d'Escherichia coli, par exemple, peuvent être alimentées en énergie et en atomes de carbone à partir du lactose disaccharide car elles sont capables de les décomposer en glucose et galactose grâce à l'enzyme bêta-galactosidase.Dans un E. coli normal qui peut contenir du lactose, il existe environ 3 000 molécules de bêta-galactosidase, soit 3 % des protéines de cette cellule ; en l'absence de lactose il n'y aura qu'une molécule de bêta-galactosidase par cellule bactérienne. La galactosidase sera synthétisée à partir de nouvelles molécules d'ARNm lorsqu'elle pourra être utilisée. Des souches mutantes d'E. coli riches en enzyme sont connues même en l'absence de lactose : ces mutants sont désavantagés par rapport aux cellules normales car ils sont contraints à une consommation inutile d'énergie et de matériaux pour produire l'enzyme qui restera sans substrat. Les substances qui provoquent une augmentation de la quantité d'enzyme, comme c'est le cas avec le lactose, seront appelées inducteurs, tandis que les enzymes seront dites inductibles. D'autres substances induisent, elles aussi de manière spécifique, la production de certaines enzymes. Toujours chez E. coli, par exemple, capable de construire tous ses acides aminés, ayant du carbone et de l'ammonium (NH3), la présence dans le milieu de culture d'un acide aminé particulier (l'histidine, par exemple) bloque la production de toutes les enzymes associées à la biosynthèse de l'acide aminé lui-même : on dira de ces enzymes qu'elles sont répressibles.Dans les cellules bactériennes les molécules d'ARNm sont détruites peu de temps après leur formation, et c'est pourquoi contrôler la production d'ARNm revient à contrôler la synthèse enzymatique en même temps temps.
L'OPÉRON
Pour expliquer comment la cellule bactérienne est capable de contrôler sa propre production d'enzymes, Jacob et Monod ont formulé l'hypothèse de l'opéron ; l'opéron est formé par plusieurs gènes qui sont fonctionnellement liés et alignés sans discontinuité le long d'un tronçon d'ADN.L'opéron se compose de trois types différents de gènes : le promoteur, où commence la formation de l'ARNm ; l'opérateur, où le contrôle est exercé; un ou plusieurs gènes de structure, qui codent pour des enzymes ou pour d'autres protéines. Dans le système bêta-galactosidase, l'opéron comprend, en plus de celui pour la bêta-galactosidase, également deux autres gènes de structure codant pour d'autres enzymes impliquées dans le métabolisme du lactose.Ces gènes sont adjacents les uns aux autres et sont transcrits les uns après les autres le long d'une même hélice d'ADN en une seule molécule d'ARNm. Les molécules d'ARNm ainsi produites sont actives pendant un temps très court, après quoi elles sont détruites par des enzymes spécifiques. .
L'activité de l'opéron est à son tour contrôlée par un autre gène, le régulateur, qui peut également être distant de l'opéron : ce régulateur code pour une protéine, appelée répresseur, qui semble se lier à l'ADN au niveau du gène opérateur. entre le promoteur et les gènes de structure bloque en fait la production d'ARNm.
Le répresseur est à son tour contrôlé, et le contrôle est effectué au moyen d'une substance "signal". Dans le cas des enzymes inductibles cette substance est "l'inducteur. L'inducteur" se lie à la molécule répresseur en modifiant sa forme pour qu'elle ne puisse plus s'adapter à l'ADN : dans ce cas, puisqu'il n'y a pas de répresseur entre le promoteur et les gènes de structure , le répresseur peut former les molécules d'ARNm et à partir de celles-ci les molécules de protéines. Avec l'épuisement de l'offre d'inducteur à nouveau le régulateur va reprendre le contrôle, ce qui arrêtera la production de nouveaux ARNm, donc de nouvelles protéines.Dans le système bêta-galactosidase l'inducteur est le lactose ou une substance très similaire à ce dérivé : ils va rejoindre le répresseur en l'inactivant pour permettre la biosynthèse des enzymes. Dans le cas des enzymes répressibles, la substance qui fait office de « signal » agit comme un corépresseur : le répresseur n'est actif que s'il est associé au corépresseur. Dans le système histidine, qui fait intervenir une dizaine d'enzymes différentes, il s'agit de cet acide aminé, associé à son ARNt, le corépresseur, l'histidine.
INTERACTIONS ALLOSTÉRIQUES
Les interactions allostériques, impliquant l'inactivation d'une enzyme en modifiant sa forme, offrent une manière différente de réguler l'activité métabolique d'une cellule. Les interactions allostériques permettent un contrôle plus précis que le système inducteur-répresseur de l'opéron, mais n'atteignent pas le résultat utile d'exclure la biosynthèse d'une substance donnée de la première étape - la production d'un ARNm.
SYSTÈMES DE CONTRLE DANS L'EUCAIRE
Certains faits portent à croire qu'un système de régulation similaire à l'opéron fonctionne et prédomine chez les plantes et les animaux. Les chromosomes de ces organismes diffèrent profondément de ceux d'E. coli et d'autres procaryotes. cellules sont très différentes.Le mécanisme de la mitose est tel que chaque cellule d'une plante ou d'un animal donné possède toutes les informations
génétique présente dans l'œuf fécondé. Par conséquent, la plupart des gènes de toute cellule spécialisée resteront inefficaces tout au long de la vie de la cellule. L'ADN de ces cellules est toujours associé à des protéines. Il est donc possible que la répression des gènes chez les eucaryotes nécessite précisément cette association entre l'ADN et les protéines.












.jpg)