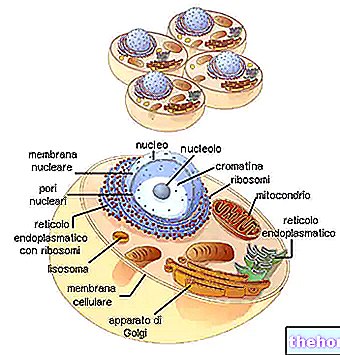
Sans variabilité génétique, tous les êtres vivants devraient (par héritage) être les mêmes que les premiers. Pour avoir des êtres inégaux, les seules explications seraient celles relatives à des créations uniques. Mais on sait que la structure de l'ADN, qui est à la base de la transmission des caractères héréditaires, a une stabilité relative et non absolue. Alors que la stabilité garantit la conservation de l'information initiale, l'instabilité détermine les modifications, ou mieux (pour employer le terme spécifique) les mutations.
Les mutations peuvent être divisées en 3 grands groupes :
- mutations génétiques ;
- mutations chromosomiques ;
- mutations génomiques.
A ce stade, il est bon d'ajouter brièvement deux concepts : l'un est celui de « réparation » et l'autre est celui de mutation « favorable » ou « défavorable ».
La réparation, terme anglais qui signifie réparation, fait référence à l'existence dans les cellules de processus enzymatiques spécifiques destinés à vérifier la conservation de l'information, en éliminant ou en corrigeant les tronçons d'ADN non conformes à l'original.
La notion de mutation « favorable » ou « défavorable » exprime la comparaison entre l'efficacité, pour l'organisme et pour l'espèce, du gène d'origine (« sauvage », c'est-à-dire « sauvage », primitif) et du gène mutant.
Il convient de noter que l'avantage et l'inconvénient font toujours référence à certaines conditions environnementales ; une mutation désavantageuse dans un certain environnement peut devenir favorable dans différentes conditions.
Les conséquences de ce principe sont vastes en génétique des populations, comme le montrent déjà les différentes efficacités de survie des variants de l'hémoglobine dans des environnements normaux ou palustres. Mais ce sujet devrait être reporté à des articles plus détaillés sur la génétique générale, humaine ou médicale.
Une autre distinction est requise (en particulier pour les mutations génétiques) entre les mutations somatiques et les mutations germinales. Toutes les cellules de notre organisme multicellulaire dérivent, comme on le sait, d'un seul zygote, mais très tôt elles se différencient en une lignée somatique, à partir de laquelle se développe tout le corps, et une lignée germinale destinée à former les gonades et, pour la méiose terminale, les gamètes Il est bien évident que, hormis les interactions entre les deux populations cellulaires, une mutation de la lignée somatique se manifestera dans un seul organisme mais n'affectera pas la descendance, alors qu'une mutation de la lignée germinale ne se produira que dans la descendance.
En ce qui concerne les différents types de mutation et les conséquences relatives, il convient de mentionner à nouveau une classification des causes de mutation. Ces causes sont appelées mutagènes, qui sont utilisées pour distinguer principalement les mutagènes physiques et chimiques. Diverses modifications de l'environnement physique peuvent conduire à des mutations, mais les principaux mutagènes physiques sont les rayonnements, c'est pourquoi les substances radioactives sont dangereuses, et ce sont surtout les particules radioactives les plus lourdes qui ont tendance à provoquer des mutations par défection, aux conséquences les plus graves.
Les mutagènes chimiques peuvent opérer à la fois en altérant la structure ordonnée des acides nucléiques, et en introduisant dans les cellules des substances similaires aux bases azotées normales, qui peuvent entrer en compétition avec ces dernières dans la synthèse des acides nucléiques, provoquant ainsi des mutations par substitution.
MUTATIONS GÉNIQUES
Les mutations génétiques affectent un ou plusieurs gènes, c'est-à-dire une portion limitée d'ADN. L'information étant stockée dans une séquence de paires de nucléotides, la plus petite unité de mutation (un muton) comprend une seule paire de bases complémentaires. Sans entrer dans le détail des différents mécanismes de mutation au niveau du gène, on peut se limiter à en citer deux : la substitution de bases et réélection (ou insertion). Dans les mutations par substitution de bases, un ou plusieurs nucléotides d'ADN sont remplacés par d'autres. Si l'erreur n'est pas corrigée à temps, une séquence suivra au moment de la transcription également altérée en ARN. Si l'altération de la triplet n'est pas limité à un synonyme (voir le code génétique), il se traduira par la substitution d'un ou plusieurs acides aminés dans la séquence polypeptidique. La substitution d'un acide aminé peut être plus ou moins critique pour la détermination de la structure de la protéine et pour sa fonction.
Dans les mutations de réélection ou d'insertion, un ou plusieurs nucléotides sont supprimés ou ajoutés à la séquence d'ADN. Ces mutations sont généralement très graves car (sauf s'il s'agit de triplets entiers qui ajoutent ou soustraient des acides aminés simples) tous les triplets qui suivent dans l'ordre de lecture sont modifiés.
Les mutations géniques sont les plus fréquentes et sont à l'origine de l'essentiel de la variabilité des traits héréditaires entre individus.
MUTATIONS CHROMOSOMIQUES
Il s'agit de mutations affectant des fragments relativement longs d'un chromosome. Il est d'usage de les classer principalement en :
- mutations chromosomiques pour réélection ;
- mutations chromosomiques par duplication ;
- mutations chromosomiques par translocation.
Les mutations dues à la défection se produisent en raison de la rupture et de la perte d'un fragment plus ou moins long de chromosome. Surtout dans la méiose, ce type de mutation est souvent mortelle, du fait de la perte totale d'un certain nombre de gènes, plus ou moins indispensables.
Dans les mutations de duplication, suite à une rupture, les moignons de chromatides ont tendance à fusionner.
Lors de l'ablation ultérieure des centromères, le chromosome, devenu dicentrique, se brise en parties souvent inégales : comme on peut le voir, il en résulte une réélection d'une part et une duplication d'autre part.
Une rupture chromosomique peut être suivie d'une inversion. Le matériel génétique total est inchangé, mais la séquence des gènes sur le chromosome est modifiée.
Le cas d'une translocation est similaire, mais il s'agit de la liaison d'un fragment d'un chromosome à un chromosome non homologue. Un chromosome est amputé et l'autre allongé ; l'information génétique totale de la cellule est toujours inchangée, mais l'effet de position est encore plus marqué. Il est facile de représenter un effet de position en se référant au concept de régulation de l'action des gènes : en changeant position sur les chromosomes, un gène peut facilement quitter un opéron et être inséré dans un autre, entraînant une "activation ou répression modifiée".
Cependant, il est dit qu'une translocation est équilibrée (ou équilibrée) lorsqu'il y a réciprocité de translocation entre deux paires de chromosomes, en gardant inchangée la somme des informations génétiques. La figure en forme de croix dans la diacinèse méiotique correspond généralement à la translocation équilibrée.
MUTATIONS GÉNOMIQUES
En rappelant que le génome est le patrimoine génétique individuel, ordonné en chromosomes, on peut préciser que l'on parle de mutations génomiques lorsque les chromosomes ont une distribution différente de la norme de l'espèce.
Les mutations génomiques peuvent être classées principalement en mutations pour la polyploïdie, l'haploïdie et l'aneuploïdie.
Les mutations de polyploïdie se produisent lorsque la réduplication n'entraîne pas de division; ils se produisent plus facilement dans les plantes, où ils sont en effet utilisés pour améliorer la production.
Si la division cellulaire échoue dans la méiose, des gamètes diploïdes peuvent apparaître; si un tel gamète parvient à fusionner avec un gamète normal, le zygote résultant de cette fécondation sera triploïde. Un tel zygote peut parfois réussir à créer un organisme entier, car la réduplication et la mitose ne nécessitent pas un nombre pair de chromosomes. Au moment de la méiose, cependant, l'appariement régulier de chromosomes homologues sera impossible.
Des mutations génomiques par haploïdie peuvent survenir lorsque, chez une espèce normalement diploïde, un gamète est activé par un autre gamète dépourvu de matière nucléaire, voire en l'absence de fécondation : un individu haploïde en résultera.
Alors que les types précédents de mutations génomiques concernent toujours l'addition ou la soustraction de nombres entiers n de chromosomes, les mutations pour aneuploïdie concernent l'excès ou le défaut de chromosomes uniques (aberrations chromosomiques).
La constitution chromosomique euploïde d'une espèce est définie comme son caryotype ou idiotype normal.
















.jpg)


