" première partie
L'enzyme qui catalyse la quatrième étape du cycle de krebs est la α-céto glutarate déshydrogénase; cette enzyme est un complexe enzymatique très similaire à la pyruvate déshydrogénase. Les deux sont composés de 48 à 60 protéines dans lesquelles trois activités enzymatiques différentes sont reconnues et ont également les mêmes cofacteurs enzymatiques ; sont des enzymes très similaires car elles agissent sur des substrats similaires : à la fois le pyruvate et le l "α-céto glutarate, sont α- les acides céto. Le mécanisme d'action des deux complexes enzymatiques est le même.
L'attaque par le pyrophosphate de thiamine sur le carbonyle (C = O) du "α-cétoglutarate, conduit à sa décarboxylation et le dérivé carboxyhydroxypropyle est formé. Avec le transfert ultérieur au lipoamide, un processus d'oxydoréduction interne a lieu, à partir duquel le dérivé carboxy du lipoamide ou le succinyl lipoamide est obtenu.
Le succinyl lipoamide réagit alors avec le coenzyme A pour donner le succinyl coenzyme A (qui se poursuit dans le cycle de krebs) et le lipoamide réduit qui est réoxydé par le FAD : le FADH2 qui se forme est réoxydé par le NAD+ et le NADH est obtenu. Dans cette étape, donc, la deuxième élimination d'un carbone du squelette carboné a eu lieu, sous forme de dioxyde de carbone.
Un groupe acyle lié à la coenzyme A est sous une forme activée, c'est-à-dire qu'il a un contenu énergétique élevé : il est donc possible d'exploiter l'énergie de la succinyl coenzyme A.

Au cinquième stade du cycle de krebs, la succinyl coenzyme A est soumise à l'action de succinyl thiokinase; deux hypothèses ont été faites sur son mode d'action : nous n'en décrirons qu'une car c'est la plus accréditée. Selon cette hypothèse, la succinyl coenzyme A est attaquée par l'azote d'une histidine (Hys) de l'enzyme : la coenzyme A est libérée et un adduit dérivé de l'histidine se forme comme intermédiaire, c'est la succinyl-enzyme (ou succinyl-Hys ) ; un orthophosphate agit sur cet intermédiaire, conduisant à la libération de succinate et à la formation de la phosphoenzyme. La phosphoenzyme, attaquée par la guanosine diphosphate (GDP), produit la guasnosine triphosphate (GTP) et l'enzyme est libérée. Du point de vue énergétique GTP = ATP : la liaison qui fournit l'énergie est la même chez les deux espèces (c'est la liaison anhydride entre le phosphoryle Β et le phosphoryle γ). Dans certains cas, le GTP est utilisé comme matériau à haute teneur énergétique mais, généralement, le GTP est converti en ATP par l'action de l'enzyme nucléoside diphospho kinase (NDPK); est une enzyme présente dans les cellules et catalyse la réaction suivante :
N1TP + N2DP → N1DP + N2TP
Nucléoside Triphosphate NiTP ® Générique
Diphosphate de nucléoside générique NiDP ®
C'est une réaction réversible; dans notre cas ça se passe :
GTP + ADP → PIB + ATP
il peut donc procéder à droite ou à gauche même pour de petites variations dans les concentrations des réactifs.
Si le cycle de krebs se déroule à une vitesse telle qu'il conduit à une production d'ATP supérieure au besoin énergétique, il y a peu d'ADP alors qu'il y a beaucoup d'ATP : la réaction catalysée par la nucléoside diphospho kinase est alors, dirigé vers la gauche (le GTP s'accumule si la nucléoside diphospho kinase n'a pas suffisamment de substrat, c'est-à-dire l'ADP). Le GTP est donc un signal de disponibilité énergétique et ralentit donc le cycle de krebs.
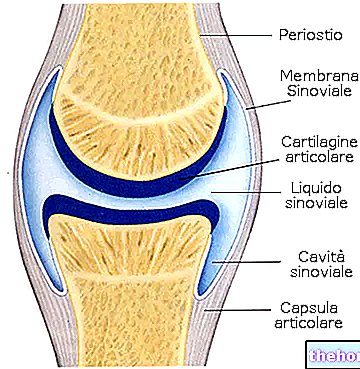
La sixième étape du cycle de krebs conduit à la formation du fumarate par l'action de succinate déshydrogénase; cette enzyme donne une réaction stéréospécifique car il se forme toujours le trans insaturé (c'est un alcène), c'est-à-dire le fumarate (alors que l'isomère cis est le maléate). La succinate déshydrogénase se trouve sur la membrane mitochondriale interne, tandis que toutes les autres enzymes du cycle de krebs sont dispersées dans la mitochondrie.
La succinate déshydrogénase a FAD comme cofacteur; il est inhibé par l'oxaloacétate (inhibition du feed-back) alors qu'il a le succinate et le fumarate comme modulateur positif (activateur). cycle de krebs nécessite de l'énergie donc la seule possibilité d'obtenir de l'oxaloacétate du patient est que la concentration du patient est très élevée : le malate est l'un des métabolites les plus concentrés dans les cellules.La réaction qui transforme le malate en oxaloacétate est également favorisée par le fait que la concentration d'oxaloacétate est maintenue faible par l'action de la citrate synthase.La réaction catalysée par la succinate déshydrogénase est donc une réaction d'auto-alimentation et c'est le seul moyen de faire se produire la transformation du malate en oxaloacétate.
La concentration en malate mitochondrial doit être compatible avec la concentration en malate cytoplasmique : ce n'est que lorsque la concentration en malate mitochondrial est si élevée qu'elle garantit la conversion du malate en oxaloacétate (dans le cycle de krebs) alors le malate peut également être utilisé dans d'autres voies (qui sont cytoplasmiques) : dans le cytoplasme le malate peut être transformé en oxaloacétate à partir duquel l'aspartate peut être obtenu par action du GOT (c'est une transaminase) ou du glucose par gluconéogenèse.
On revient à la septième étape du cycle de krebs elle est catalysée par l'enzyme fumerasi: de l'eau est ajoutée de manière stéréospécifique pour fabriquer du L-malate.
Dans la dernière étape du cycle de Krebs, dont nous avons déjà parlé, l'action du malate déshydrogénase. Cette enzyme utilise une molécule NAD+ pour son action catalytique.
Nous avons ainsi conclu la description des différentes étapes du cycle de krebs.
Le cycle de krebs est totalement réversible.
Pour augmenter la vitesse du cycle de krebs, la concentration des métabolites présents dans ce cycle peut être augmentée ; une des stratégies pour augmenter la vitesse du cycle de krebs consiste à convertir une partie du pyruvate qui entre dans les mitochondries en oxaloacétate (par l'action de la pyruvate carboxylase) et de ne pas tout transformer en acétyl coenzyme A : augmente ainsi la concentration en oxaloacétate qui est un métabolite du cycle de krebs et, par conséquent, augmente la vitesse de l'ensemble du cycle.
Dans le cycle de krebs, trois NAD+ sont convertis en trois NADH et un FAD en FADH2 et, de plus, un GTP est obtenu : en canalisant le pouvoir réducteur obtenu à partir du cycle de krebs, de l'ATP supplémentaire est produit ; dans la chaîne respiratoire, le pouvoir réducteur est transféré du NADH et du FADH2 à l'oxygène : ce transfert est dû à une série d'enzymes situées sur la membrane mitochondriale qui, dans leur action, conduisent à la production d'ATP.
Les processus de la chaîne respiratoire sont des processus exergoniques et l'énergie libérée est utilisée pour produire de l'ATP ; le but de la cellule est d'exploiter les processus exergoniques pour faire en sorte que la synthèse de l'ATP ait lieu. Pour chaque molécule de NADH qui entre dans la chaîne respiratoire, on obtient 2,5 molécules d'ATP et pour chaque FADH2 1,5 molécules d'ATP sont obtenues ; cette diversité est due au fait que FADH2 entre dans la chaîne respiratoire à un niveau inférieur à celui du NADH.
Avec le pouvoir réducteur du métabolisme aérobie, 30-32 ATP (219-233 kcal / mol) sont obtenus avec une efficacité d'environ 33% (l'efficacité du métabolisme anaérobie est d'environ 2%).