Définition
Les enzymes sont des protéines produites dans les cellules végétales et animales, qui agissent comme des catalyseurs accélérant les réactions biologiques sans être modifiées.
Les enzymes fonctionnent en se combinant avec une substance spécifique pour la transformer en une substance différente ; des exemples classiques sont donnés par les enzymes digestives présentes dans la salive, l'estomac, le pancréas et l'intestin grêle, qui remplissent une fonction essentielle dans la digestion et aident à décomposer les aliments en constituants de base, qui peuvent ensuite être absorbés et utilisés par le corps, transformés par d'autres enzymes ou excrété comme déchet.
Chaque enzyme a un rôle spécifique : celui qui décompose les graisses, par exemple, n'agit pas sur les protéines ou les glucides. Les enzymes sont indispensables au bien-être de l'organisme, la carence, même d'une seule enzyme, peut entraîner de graves problèmes.Un exemple bien connu est la phénylcétonurie (PCU), une maladie caractérisée par l'incapacité à métaboliser un acide aminé essentiel, la phénylalanine, dont l'accumulation peut provoquer des malformations physiques et des maladies mentales.
Étude biochimique
Les enzymes sont des protéines particulières qui ont la caractéristique d'être des catalyseurs biologiques, c'est-à-dire qu'elles ont la capacité de décomposer l'énergie d'activation (Eatt) d'une réaction, en modifiant son cheminement de sorte qu'un processus cinétiquement lent soit plus rapide.
Les enzymes augmentent la cinétique des réactions thermodynamiquement possibles et, contrairement aux catalyseurs, elles sont plus ou moins spécifiques : elles possèdent donc une spécificité de substrat.
L'enzyme n'intervient pas dans la stoechiométrie de la réaction : pour cela, le site catalytique final doit être identique à celui de départ.
Dans l'action catalytique, il y a presque toujours une phase lente qui détermine la vitesse du processus.
Quand on parle d'enzymes, il n'est pas correct de parler de réactions d'équilibre, on parle plutôt de régime permanent (état dans lequel un certain métabolite est formé et consommé en continu, maintenant sa concentration presque constante dans le temps). Le produit d'une réaction catalysée par une enzyme est généralement lui-même un réactif pour une réaction ultérieure, catalysé par une autre enzyme, et ainsi de suite.
Les processus catalysés par des enzymes consistent généralement en des séquences de réactions.
Une réaction générique catalysée par une enzyme (E) peut être résumée comme suit :

E est l'enzyme
S est le substrat ;
ES représente le produit d'addition entre l'enzyme et le substrat ;
P est le produit ;
K est la constante de vitesse de la réaction.
Une enzyme générique (E) se combine avec le substrat (S) pour former l'adduit (ES) avec une constante de vitesse K1 ; il peut se dissocier en E + S, avec une constante de vitesse K2, ou, (si « vit » assez longtemps ) peut procéder à la forme P avec une constante de vitesse K3.
Le produit (P) peut, à son tour, se recombiner avec l'enzyme et reformer l'adduit avec une constante de vitesse K4.
Lorsque l'enzyme et le substrat sont mélangés, il y a une fraction de temps pendant laquelle la rencontre entre les deux espèces n'a pas encore eu lieu : c'est-à-dire qu'il y a un intervalle de temps extrêmement court (qui dépend de la réaction) dans lequel l'enzyme et le substrat ont pas encore atteint ; après cette période, l'enzyme et le substrat entrent en contact en quantités croissantes et l'adduit ES est formé. Par la suite, l'enzyme agit sur le substrat et le produit est libéré. On peut alors dire que c" est un intervalle de temps initial dans lequel la concentration de l'adduit ES ne peut être définie ; après cette période, on suppose qu'un état stationnaire est établie, c'est-à-dire que la vitesse des processus qui conduisent à l'obtention de l'adduit est égale à la vitesse des processus qui conduisent à la destruction de l'adduit.
La constante de Michaelis-Menten (KM) est une constante d'équilibre (référée au premier équilibre décrit ci-dessus) ; on peut dire, avec une bonne approximation (car K3 devrait également être considéré), que KM est représenté par le rapport entre les constantes cinétiques K2 et K1 (en référence à la destruction et la formation de l'adduit ES dans le premier équilibre décrit ci-dessus) .
Grâce à la constante de Michaelis-Menten, nous avons une « indication de l'affinité entre l'enzyme et le substrat : si le KM est petit c » est une « affinité élevée entre l'enzyme et le substrat, alors l'adduit ES est stable.
Les enzymes sont soumises à une régulation (ou modulation).
Autrefois on parlait surtout de modulation négative, c'est-à-dire d'inhibition des capacités catalytiques d'une enzyme mais, il peut aussi y avoir une modulation positive, c'est-à-dire qu'il existe des espèces capables d'augmenter les capacités catalytiques d'une enzyme.
Il existe 4 types d'inhibitions (obtenues à partir d'approximations faites sur un modèle pour faire correspondre des données expérimentales avec des équations mathématiques) :
- inhibition compétitive
- inhibition non compétitive
- inhibition incompétitive
- inhibition compétitive
On parle d'inhibition compétitive lorsqu'une molécule (inhibiteur) est capable de rivaliser avec le substrat. Pour la similarité structurelle, l'inhibiteur peut réagir à la place du substrat, d'où la terminologie "inhibition compétitive".La probabilité que l'enzyme se lie à l'inhibiteur ou au substrat dépend de la concentration des deux et de leur affinité avec l'enzyme ; la vitesse de réaction dépend donc de ces facteurs.
Pour obtenir la même vitesse de réaction que sans la présence de l'inhibiteur, il est nécessaire d'avoir une concentration en substrat plus élevée.
Il est montré expérimentalement qu'en présence d'un inhibiteur, la constante de Michaelis-Menten augmente.
En ce qui concerne, au contraire, "l'inhibition non compétitive, l'interaction entre la molécule qui devrait fonctionner comme un modulateur (positif ou négatif-inhibiteur) et "l'enzyme, a lieu dans un site différent de celui dans lequel l'interaction se produit entre l'enzyme et le substrat; on parle donc de modulation allostérique (du grec allostères → autre site).
Si l'inhibiteur se lie à l'enzyme, il peut induire un changement dans la structure de l'enzyme et, par conséquent, peut diminuer l'efficacité avec laquelle le substrat se lie à l'enzyme.
Dans ce type de procédé, la constante de Michaelis-Menten reste constante puisque cette valeur dépend des équilibres entre l'enzyme et le substrat et, même en présence d'un inhibiteur, ces équilibres ne changent pas.
Le phénomène d'inhibition incompétitive est rare ; un inhibiteur incompétitif typique est une substance qui se lie de manière réversible à l'adduit ES donnant lieu à l'ESI :

L'inhibition de l'excès de substrat peut parfois être incompétitive, car cela se produit lorsqu'une seconde molécule de substrat se lie au complexe ES, donnant naissance au complexe ESS.
Un inhibiteur compétitif, en revanche, ne peut se lier qu'à l'adduit enzymatique substrat comme dans le cas précédent : la liaison du substrat à l'enzyme libre induit une modification conformationnelle qui rend le site accessible pour l'inhibiteur.
La constante de Michaelis Menten diminue à mesure que la concentration en inhibiteur augmente : apparemment, donc, l'affinité de l'enzyme pour le substrat augmente.
Sérine protéase
Il s'agit d'une famille d'enzymes à laquelle appartiennent la chymotrypsine et la trypsine.
La chymotrypsine est une enzyme protéolytique et hydrolytique qui coupe à droite les acides aminés hydrophobes et aromatiques.
Le produit du gène qui code pour la chymotrypsine n'est pas actif (il est activé avec une commande) ; la forme inactive de la chymotrypsine est représentée par une chaîne polypeptidique de 245 acides aminés. La chymotrypsine a une forme globulaire due à cinq ponts disulfures et à d'autres interactions mineures (électrostatique, forces de Van der Waals, liaisons hydrogène, etc.).
La chymotrypsine est produite par les cellules chymosées du pancréas où elle est contenue dans des membranes spéciales et expulsée par le canal pancréatique dans l'intestin, au moment de la digestion des aliments : la chymotrypsine est en fait une enzyme digestive. Les protéines et les nutriments que nous ingérons par l'alimentation sont soumis à une digestion pour être réduits en chaînes plus petites et être absorbés et transformés en énergie (par exemple, les amylases et les protéases décomposent les nutriments en glucose et en acides aminés qui atteignent les cellules, à travers les vaisseaux sanguins ils atteignent la veine porte et de là sont transportés vers le foie où ils subissent un traitement supplémentaire).
Les enzymes sont produites sous une forme non active et ne sont activées que lorsqu'elles atteignent le « site où elles doivent opérer » ; une fois leur action terminée, ils sont désactivés. Une enzyme, une fois désactivée, ne peut pas être réactivée : pour avoir une « action catalytique supplémentaire, elle doit être remplacée par » une autre molécule d'enzyme. Si la chimitrypsine était produite sous forme active déjà dans le pancréas, elle s'attaquerait à ce dernier : les pancréatites sont des pathologies dues à des enzymes digestives déjà activées dans le pancréas (et non dans les sites requis) ; certaines d'entre elles si elles ne sont pas traitées à temps, conduire à la mort.
Dans la chymotrypsine et dans toutes les protéases à sérine, l'action catalytique est due à l'existence de l'anion alcool (-CH2O-) dans la chaîne latérale d'une sérine.
Les protéases à sérine prennent ce nom précisément parce que leur action catalytique est due à une sérine.
Une fois que toute l'enzyme a effectué son action, avant de pouvoir à nouveau opérer sur le substrat, il faut la restituer avec de l'eau ; la "libération" de la sérine par l'eau est l'étape la plus lente du processus, et c'est cette phase qui détermine la vitesse de catalyse.
L'action catalytique se déroule en deux phases :
- formation de l'anion aux propriétés catalytiques (alcoolate d'anion) et attaque nucléophile subséquente sur le carbone carbonyle (C = O) avec clivage de la liaison peptidique et formation de l'ester ;
- attaque par l'eau avec restauration du catalyseur (capable d'exercer à nouveau son action catalytique).
Les différentes enzymes appartenant à la famille des sérine protéases peuvent être constituées de différents acides aminés mais, pour toutes, le site catalytique est représenté par l'anion alcoolique de la chaîne latérale d'une sérine.
Une sous-famille de sérine protéases est celle des enzymes impliquées dans la coagulation (qui consiste en la transformation des protéines, de leur forme inactive en une « autre forme active). Ces enzymes assurent une coagulation la plus efficace possible et limitée en l'espace et le temps (la coagulation doit se produire rapidement et ne doit se produire qu'au voisinage de la zone lésée). Les enzymes impliquées dans la coagulation sont activées en cascade (à partir de l'activation d'une seule enzyme, des milliards d'enzymes sont obtenues : chaque enzyme activée , active à son tour de nombreuses autres enzymes).
La thrombose est une pathologie due au dysfonctionnement des enzymes de la coagulation : elle est provoquée par l'activation, sans nécessité (car il n'y a pas de lésion), des enzymes utilisées dans la coagulation.
Il existe des enzymes modulatrices (régulatrices) et des enzymes inhibitrices pour d'autres enzymes : en interagissant avec ces dernières, elles régulent ou inhibent leur activité ; même le produit d'une enzyme peut être un inhibiteur de l'enzyme.Il y a aussi des enzymes qui fonctionnent plus, plus le substrat présent est important.
Lysozyme
Luigi Pasteur a découvert, en éternuant sur une boîte de Pétri, que dans le mucus se trouve une enzyme capable de tuer les bactéries : lysozyme; du grec : liso = quelle taille ; zimo = enzyme.
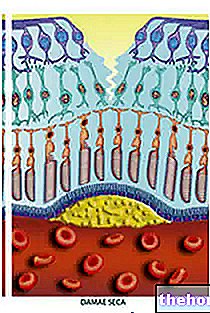
Le lysozyme est capable de briser la paroi cellulaire des bactéries. Les bactéries, et les organismes unicellulaires en général, ont besoin de structures mécaniquement résistantes qui limitent leur forme ; à l'intérieur des bactéries, il y a une pression osmotique très élevée, de sorte qu'elles attirent l'eau. La membrane plasmique exploserait s'il n'y avait pas de paroi cellulaire qui s'oppose à l'entrée d'eau et limite le volume de la bactérie.
La paroi cellulaire est constituée d'une chaîne polysaccharidique dans laquelle alternent des molécules de N-acétyl-glucosamine (NAG) et des molécules d'acide N-acétyl-muramique (NAM) ; la liaison entre NAG et NAM est rompue par hydrolyse. Le groupe carboxyle de NAM, dans la paroi cellulaire, est engagé dans une liaison peptidique avec un acide aminé.
Entre les différentes chaînes, se forment des ponts constitués de liaisons pseudo-peptidiques : la ramification est due à la molécule de lysine ; la structure dans son ensemble est très ramifiée et cela lui confère une grande stabilité.
Le lysozyme est un antibiotique (tue les bactéries) : il agit en créant une fissure dans la paroi bactérienne ; lorsque cette structure (qui est mécaniquement résistante) se brise, la bactérie puise de l'eau jusqu'à ce qu'elle éclate. Le lysozyme parvient à rompre la liaison glucosidique -1,4 entre NAM et NAG.
Le site catalytique du lysozyme est représenté par un sillon qui longe l'enzyme dans lequel la chaîne polysaccharidique est insérée : six anneaux glucosidiques de la chaîne sont placés dans le sillon.
En position trois de la rainure c" est un starter : dans cette position un seul NAG peut être placé, car le NAM, qui est de dimensions supérieures, ne peut pas entrer. Le site catalytique réel se situe entre les positions quatre et cinq : puisqu'il y a un NAG en position trois, la coupure se fera entre un NAM et un NAG (et non l'inverse) ; la coupure est donc spécifique.
Le pH optimal pour que le lysozyme fonctionne est de cinq. Dans le site catalytique de l'enzyme, c'est-à-dire entre les positions quatre et cinq, se trouvent les chaînes latérales d'un acide aspartique et d'un acide glutamique.
Degré d'homologie : mesure la parenté (c'est-à-dire la similitude) entre les structures protéiques.
Il existe une forte relation entre le lysozyme et la lactose-synthase.
La lactose synthétase synthétise le lactose (qui est le principal sucre du lait) : le lactose est un galactosyl glucoside dans lequel c"est une liaison -1,4 glucosidique entre le galactose et le glucose.
Par conséquent, la lactose synthétase catalyse la réaction opposée à celle catalysée par le lysozyme (qui divise à la place la liaison glucosidique -1,4).
La lactose synthétase est un dimère, c'est-à-dire qu'elle est constituée de deux chaînes protéiques, dont l'une a des propriétés catalytiques et est comparable au lysozyme et l'autre est une sous-unité régulatrice.
Pendant la grossesse, les glycoprotéines sont synthétisées par les cellules de la glande mammaire par l'action de la galatosyl-transférase (elle a une « homologie de séquence de 40 % avec le lysozyme) : cette enzyme est capable de transférer un groupement galactosyle d'une structure à haute énergie à une structure glycoprotéique Pendant la grossesse, l'expression du gène qui code pour la galactosisyl-transférase est induite (il y a aussi l'expression d'autres gènes qui donnent aussi d'autres produits) : il y a une augmentation de la taille du sein car il est activé la glande mammaire (auparavant inactive) qui doit produire du lait Lors de l'accouchement, il se produit de la -lactalbumine qui est une protéine régulatrice : elle est capable de réguler la capacité catalytique de la galactosyl-transférase (par discrimination du substrat). La galactosyl-transférase modifiée par la -lactalalbumine est capable de transférer un galactosyle sur une molécule de glucose : formant une liaison -1,4 glycosidique et donnant du lactose (lactose synthétase).
Par conséquent, la galactose transférase prépare la glande mammaire avant l'accouchement et produit du lait après l'accouchement.
Pour produire des glycoprotéines, la galactosyl transférase se lie à un galactosyl et à un NAG ; lors de l'accouchement, l'albumine lactale se lie à la galactosyltransférase faisant que cette dernière reconnaît le glucose et non plus le NAG pour donner du lactose.
.jpg)